Lapwing

Introduction
The noisy, tumbling display flights of this green and white plover are a welcome, if increasingly uncommon, sign of spring.
The Lapwing is a handsome wader with its glossy dark green plumage (often looking black from a distance) and long crest. It used to breed commonly over much of lowland Britain, but atlas projects have charted a reduction in the number of breeding locations, caused by changes in land management and increased predation pressure.
In winter, the Lapwing is widely distributed in lowland Britain, but BTO Surveys have shown that numbers have both shifted eastward (perhaps a response to milder winters) and increased in coastal wetlands as birds appear to feed more on mudflats at low tide.
- Our Trends Explorer gives you the latest insight into how this species' population is changing.
Key Stats

Identification
Songs and Calls
Song:
Alarm call:
Status and Trends
Conservation Status
Population Change
Although CBC recorded some increase in its early years, Lapwings have declined continuously on lowland farmland since the mid 1980s. National surveys in England and Wales showed a 49% population decline between 1987 and 1998 (Wilson et al. 2001). In Northern Ireland, the breeding population had shrunk to just 860 (277-1545) pairs by 2013, representing a decrease of around 89% since 1987, with the distribution becoming increasingly fragmented (Colhoun et al. 2015). Population declines there mirror similar declines throughout wet meadow areas of Wales and southeast England (Wilson et al. 2001, 2005a). The BBS map of change in relative density between 1994-96 and 2007-09 indicates that decreases have been strongest in lowland regions and the south and that some increase may have occurred in some upland and northern regions of Britain. Stability in breeding numbers in the Uists between 1983 and 2014 runs against the UK trend and heightens the relative importance of this population (Calladine et al. 2015). Winter numbers counted by WeBS, mainly at coastal sites and omitting some big concentrations inland, increased in Britain during the 1980s and early 1990s, but decreased steeply between 2005 and 2010 (Frost et al. 2020); these birds are mainly of continental origin. Lapwing is in decline across Europe, having decreased in all regions since 1980, although with differing regional timing (PECBMS: PECBMS 2009, PECBMS 2020a>). The 2009 review moved this species from amber to the UK red list, for which it continues to qualify on the strength of its UK decline.
Distribution
In Britain, wintering concentrations of Lapwings are highest in the Somerset Levels, Fens, northwest England, the Uists, Caithness and Orkney. The former core area in central England is less dominant and Fenland and the east coast now more prominent than in the 1981–84 Winter Atlas. Densities of breeding Lapwings are highest in northwest England, Orkney, Shetland and the Outer Hebrides; those in Ireland are low throughout.
Occupied 10-km squares in UK
or view it on Bird Atlas Mapstore.
or view it on Bird Atlas Mapstore.
European Distribution Map
Distribution Change
Despite decades of population decline, Lapwings are still the most widespread breeding waders in Britain & Ireland. Over the last 40 years, they have been lost as breeders from most of southwest England, west Wales and western mainland Scotland. Losses throughout Ireland mean that the current range is less than half that in the 1968–72 Breeding Atlas.
Change in occupied 10-km squares in the UK
or view it on Bird Atlas Mapstore.
or view it on Bird Atlas Mapstore.
Seasonality
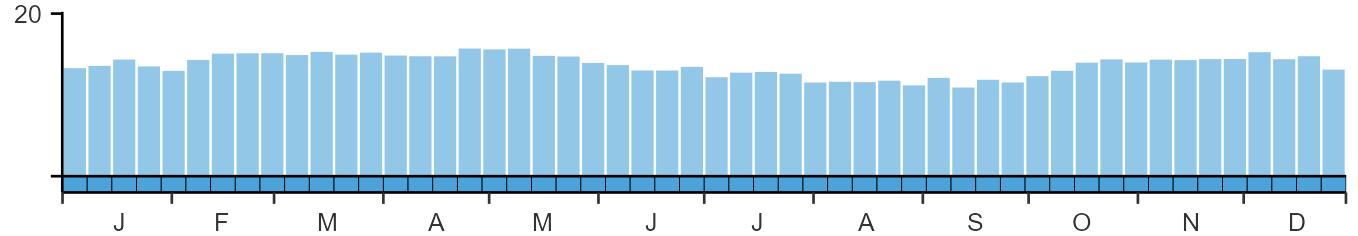
Lapwings are widely reported and present year-round.
Weekly pattern of occurrence
The graph shows when the species is present in the UK, with taller bars indicating a higher likelihood of encountering the species in appropriate regions and habitats.
Habitats
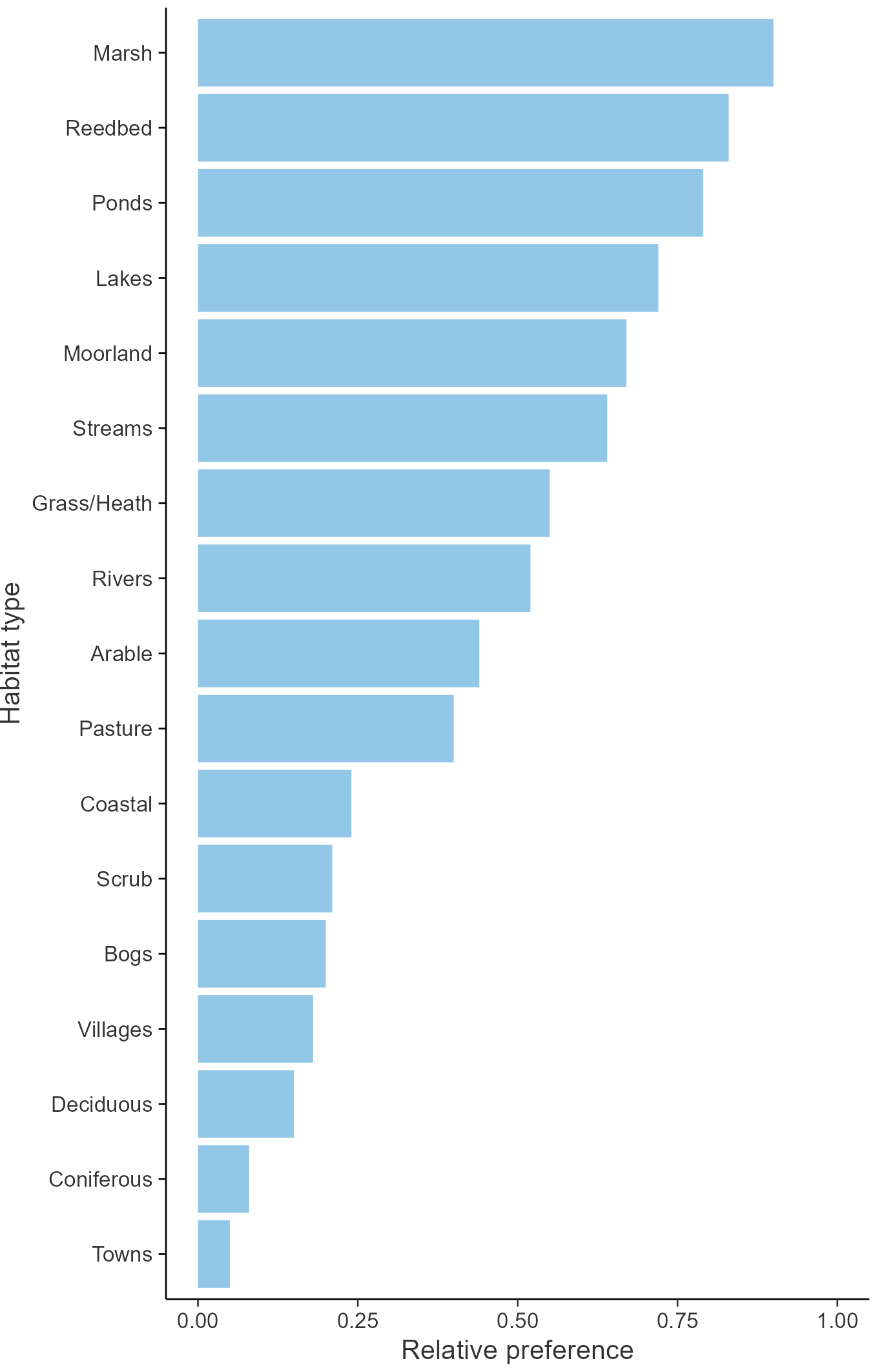
Breeding season habitats
Relative frequency by habitat
The graph shows the habitats occupied in the breeding season, with the most utilised habitats shown at the top. Bars of similar size indicate the species is equally likely to be recorded in those habitats.
Movement
Britain & Ireland movement
Foreign locations of birds ringed or recovered in Britain & Ireland
Dots show the foreign destinations of birds ringed in Britain & Ireland, and the origins of birds ringed overseas that were subsequently recaptured, resighted or found dead in Britain & Ireland. Dot colours indicate the time of year that the species was present at the location.
- Winter (Nov-Feb)
- Spring (Mar-Apr)
- Summer (May-Jul)
- Autumn (Aug-Oct)

European movements
EuroBirdPortal uses birdwatcher's records, such as those logged in BirdTrack to map the flows of birds as they arrive and depart Europe. See maps for this species here.
The Eurasian-African Migration Atlas shows movements of individual birds ringed or recovered in Europe. See maps for this species here.
Biology
Productivity and Nesting
Nesting timing
Egg measurements
Clutch Size
Survival and Longevity
Survival is shown as the proportion of birds surviving from one year to the next and is derived from bird ringing data. It can also be used to estimate how long birds typically live.
View number ringed each year in the Online Ringing Report.
lifespan
Survival of adults
Survival of juveniles
Biometrics
Wing length and body weights are from live birds (source).
Wing length
Body weight
Ring Size
Classification, names and codes
Classification and Codes
- Order: Charadriiformes
- Family: Charadriidae
- Scientific name: Vanellus vanellus
- Authority: Linnaeus, 1758
- BTO 2-letter code: L.
- BTO 5-letter code: LAPWI
- Euring code number: 4930
Alternate species names
- Catalan: fredeluga europea
- Czech: cejka chocholatá
- Danish: Vibe
- Dutch: Kievit
- Estonian: kiivitaja
- Finnish: töyhtöhyyppä
- French: Vanneau huppé
- Gaelic: Curracag
- German: Kiebitz
- Hungarian: bíbic
- Icelandic: Vepja
- Irish: Pilibín
- Italian: Pavoncella
- Latvian: kivite
- Lithuanian: paprastoji pempe
- Norwegian: Vipe
- Polish: czajka (zwyczajna)
- Portuguese: abibe
- Slovak: cíbik chochlatý
- Slovenian: priba
- Spanish: Avefría europea
- Swedish: tofsvipa
- Welsh: Cornchwiglen
- English folkname(s): Peewit, Green Plover, Hornpie, Flopwing
Research
Causes of Change and Solutions
Causes of change
There is good evidence that declines have resulted from habitat loss and degradation due to changes in agricultural practice, in particular change from spring to autumn sowing, drainage of grasslands and loss of mixed farmland, which have led to breeding productivity dropping below a sustainable level. Chick mortality is thought to be the main determinant of poor Lapwing productivity, and therefore of population decline.
Further information on causes of change
The decline of the Lapwing exemplifies how different factors can combine to cause population change (Robinson et al. 2014). The decrease in the 1980s was due to a period of low survival, when annual life expectancy decreased from 7 years to just 4.5 years, caused by a series of cold winters. At the same time, though, average nest survival decreased markedly, meaning the population could not recover from what would normally have been only a temporary setback.
There is a good deal of research supporting the hypothesis that habitat loss and degradation due to the intensification of farming have reduced breeding productivity (e.g. Galbraith 1988, Shrubb 1990, Hotker 1991, Hudson et al. 1994, Siriwardena et al. 2000a, Taylor & Grant 2004, Wilson et al. 2005, Milsom 2005, Fuller & Ausden 2008). These changes include extensive drainage, increased use of pesticides and fertilisers, re-seeding, earlier and more frequent mowing, increased grazing pressure and loss of spring cereals. Increases in intensity of grazing have reduced the habitat quality for Lapwing (Shrubb 1990, Fuller & Ausden 2008), whilst fertilisation has led to earlier spring grass growth, earlier cutting dates and higher stocking levels, which have increased egg and chick mortality and reduced relaying opportunities (Durant et al. 2008). Drainage and loss of wet features on grassland have also had a negative impact, reducing food supplies (Taylor & Grant 2004, Eglington et al. 2010).
Loss of mixed farming systems and extensive grazing have reduced the availability of high-quality foraging habitat close to nesting habitat, i.e. unimproved pasture and meadows, to birds breeding in arable areas, resulting in reduced breeding success (Galbraith 1988, Hudson et al. 1994, Henderson et al. 2004).
In the uplands, afforestation has also resulted in habitat loss (Fuller & Ausden 2008). It is possible that afforestation may also cause indirect as well as direct habitat loss: a study in Estonia found that breeding sites were preferentially located further away from woodland and suggested that woodland planting may have contributed to declines there (Kaasiku et al. 2019); however, similar analysis has not been carried out in the UK. On arable land, spring-sown cereals were once favoured nesting crops but these have been widely replaced by autumn-sown cereals, which are less suitable breeding habitats (Shrubb 1990, Shrubb et al. 1991, Mason & Macdonald 1999, Fuller & Ausden 2008). Land use changes causing a reduction in spring sward height also probably contributed to a decline on mixed farmland habitat in Scotland (Bell & Calladine 2017).
Lapwing population declines may also be explained partly by increased nest predation rates resulting from habitat changes due to agricultural intensification (Baines 1990, Liker & Szekely 1997, Jackson & Green 2000, Chamberlain & Crick 2003, Evans 2004, Jackson et al. 2004, Milsom 2005, Bolton et al. 2007, Teunissen et al. 2008, MacDonald & Bolton 2008b, Bellebaum & Bock 2009, Mason et al. 2018). Long-term nest record card analysis has shown that the proportion of nests lost to predators was substantially higher in the 1990s than in previous decades (Sharpe et al. 2008).
Recent empirical evidence suggests that levels of predation on wader nests are unsustainably high in many cases, even in some situations where breeding habitat is otherwise favourable (MacDonald & Bolton 2008a). Laidlaw et al. (2015, 2017) found that nest predation rates in wet grassland increased as the distance from patches of taller vegetation increased, and suggested that the distribution and activity of predators might be affected by the vegetation. Small mammal activity was higher in tall vegetation, particularly later in the breeding season, hence foxes in the vicinity of vegetation patches may be attracted to them and away from Lapwing nests and chicks (Laidlaw et al. 2018). In dry fields, nest predation rates were higher further from field edges (Laidlaw et al. 2017). Predation rates are also higher in areas with low Lapwing density (Laidlaw et al. 2017).
In the Uists, where the overall population is stable (Calladine et al. 2015), clutch survival is significantly lower in areas where introduced Hedgehogs Erinaceus europaeus are more abundant; however the impact of predation on local populations was unclear and more complex factors may influence trends (Calladine et al. 2017).
A Danish study found a negative correlation between annual predation rates and the numbers of breeding Lapwing, suggesting that birds nesting at higher densities may be more efficient at defending against predators (Moller et al. 2018) and hence that declines could make populations even more vulnerable if they also lead to lower nesting densities. McCallum et al. (2015, 2016) found that Lapwing density was greatest at higher elevation, but only where soils were less peaty and less acidic, opening the way to trials of whether soil amendments such as liming could contribute to conservation management for breeding Lapwings and other species of concern that depend upon soil-dwelling invertebrates. Declines among Lapwings are unlikely to be ameliorated by either habitat improvement or predator control in isolation, however (Bodey et al. 2011, Smart et al. 2013).
Information about conservation actions
As a result of the declines, the Lapwing has been the subject of a substantial amount of recent research and a number of conservation actions have been suggested. Local management actions to improve breeding habitat could include the creation of Lapwing plots in arable fields (Schmidt et al. 2017); sowing spring cereals rather than autumn cereals; delaying mowing or grazing; management of water levels to restore wetland features and habitats (Eglington et al. 2010); the exclusion of livestock from selected areas on coastal grazing marshes where the rate of grass growth is slow in spring (Hart et al. 2002); and soil amendment on upland farmland (McCallum et al. 2015).
The exclusion of predators such as foxes may also be important (see Causes of Change section), and can improve nest success (Sheldon et al. 2007; Rickenbach et al. 2011; Malpas et al. 2013b), although one study found that predator control did not affect nesting success (Bodey et al. 2011). An alternative, and possibly more sustainable, approach could be to influence predation rates by the creation and management of habitat to influence the movements of predators away from nests; for example taller vegetation patches which attract predators as a result of higher vole numbers, and wetland features to deter some predators (Laidlaw et al. 2015; 2017; 2018).
However, declines among Lapwings are unlikely to be ameliorated by either habitat improvement or predator control in isolation (Bodey et al. 2011, Smart et al. 2013 Smart et al. 2013) and hence an approach which combines different conservation actions across sites and across wider landscapes may be required. An Estonian study found that apparently suitable sites close to woodland were avoided (Kaasiku et al. 2019); hence habitat improvement should be focused on areas away from woodland and new woodlands should not be planted close to existing sites.
Publications (8)
Watching Out for Waders: The Working for Waders Nest Camera Project
Author: Noyes, P., Laurie, P., Wetherhill, A. & Wilson, M.
Published: 2024
This report presents the results of a trial involving the use of trail cameras by land managers and other wader conservation stakeholders to monitor the outcome of wader nesting attempts. It presents the results of the trial and assesses the potential for the project to improve wader conservation knowledge and management.
04.10.24
Reports Research reports

Loss of breeding waders from key lowland grassland sites in Northern Ireland
Author: Booth Jones, K.A., O’Connell, P., Wolsey, S., Carrington-Cotton, A., Noble, D.G., McCulloch, N. & Calladine, J.R.
Published: 2022
Between the mid-1980s and 2018–2019, Northern Ireland’s lowland wet grasslands saw a 73% decline in their breeding wader populations, from 1,296 to 354 pairs across 74 surveyed sites.
18.07.22
Papers

Nesting dates of Moorland Birds in the English, Welsh and Scottish Uplands
Author: Wilson, M.W., Fletcher, K., Ludwig, S.C. & Leech, D.I.
Published: 2022
Rotational burning of vegetation is a common form of land management in UK upland habitats, and is restricted to the colder half of the year, with the time period during which burning may be carried out in upland areas varying between countries. In England and Scotland, this period runs from the 1st October to 15th April, but in the latter jurisdiction, permission can be granted to extend the burning season to 30th April. In Wales, this period runs from 1st October to 31st March.This report sets out timing of breeding information for upland birds in England, Scotland and Wales, to assess whether rotational burning poses a threat to populations of these species, and the extent to which any such threat varies in space and time.
17.02.22
Reports Research reports

Sensitivity mapping for breeding waders in Britain: towards producing zonal maps to guide wader conservation, forest expansion and other land-use changes. Report with specific data for Northumberland and north-east Cumbria
Author: O’Connell, P., Wilson, M., Wetherhill, A. & Calladine, J.
Published: 2021
Breeding waders in Britain are high profile species of conservation concern because of their declining populations and the international significance of some of their populations. Forest expansion is one of the most important, ongoing and large-scale changes in land use that can provide conservation and wider environmental benefits, but also adversely affect populations of breeding waders. We describe models to be used towards the development of tools to guide, inform and minimise conflict between wader conservation and forest expansion.Extensive data on breeding wader occurrence is typically available at spatial scales that are too coarse to best inform waderconservation and forestry stakeholders. Using statistical models (random forest regression trees) we model the predicted relative abundances of 10 species of breeding wader across Britain at 1-km square resolution. Bird data are taken from Bird Atlas 2007–11, which was a joint project between BTO, BirdWatch Ireland and the Scottish Ornithologists’ Club, and modelled with a range of environmental data sets.
09.12.21
Reports Research reports

Monitoring Breeding Waders in Wensleydale: trialling surveys carried out by farmers and gamekeepers
Author: Jarrett, D., Calladine, J., Wernham, C., Wilson, M.
Published: 2017
21.11.17
Reports
Consequences of population change for local abundance and site occupancy of wintering waterbirds
Author: Méndez, V., Gill, J.A., Alves, J.A., Burton, N.H.K. & Davies, R.G.
Published: 2017
Protected sites for birds are typically designated based on the site’s importance for the species that use it. For example, sites may be selected as Special Protection Areas (under the European Union Directive on the Conservation of Wild Birds) if they support more than 1% of a given national or international population of a species or an assemblage of over 20,000 waterbirds or seabirds. However, through the impacts of changing climates, habitat loss and invasive species, the way species use sites may change. As populations increase, abundance at existing sites may go up or new sites may be colonized. Similarly, as populations decrease, abundance at occupied sites may go down, or some sites may be abandoned. Determining how bird populations are spread across protected sites, and how changes in populations may affect this, is essential to making sure that they remain protected in the future.
20.09.17
Papers

The decline of a population of farmland breeding waders: a twenty-five-year case study
Author: M.V. Bell, Calladine, J.
Published: 2017
The breeding populations of many different wader species are in decline across the globe, and the UK is no exception. These declines have been linked to increased predator numbers, changes in agricultural practices, and in the management of the wider landscape. There is an urgent need for information on how such changes in land management, particularly within farmland, may affect breeding waders. This information can then be used to inform future land management decisions.
12.04.17
Papers Bird Study

Distribution shifts in wintering Golden Plover Pluvialis apricaria and Lapwing Vanellus vanellus in Britain
Author: Gillings, S., Austin, G.E., Fuller, R.J. & Sutherland, W.J.
Published: 2006
01.01.06
Papers Bird Study



